Objective To investigate the molecular mechanism of luteolin-regulated epithelial-mesenchymal transition(EMT) in non-small cell lung cancer. Methods Firstly,the potential target of luteolin was studied by molecular docking.The effect of luteolin on EMT markers was analyzed,and then molecular biology experiments were used to verify the results. Results The result of molecular docking showed that luteolin had a good docking effect on the integrin family,of which the lowest binding energy was -15.11 in docking with Integrin αIIbβ3.The result of flow cytometry showed that luteolin could down-regulate the expression of integrin induced by hypoxia. Conclusion Luteolin can regulate integrin expression ,which suggested that luteolin can regulate EMT through integrin.This conclusion may provide new methods in prevention of tumor metastasis for traditional Chinese medicine.
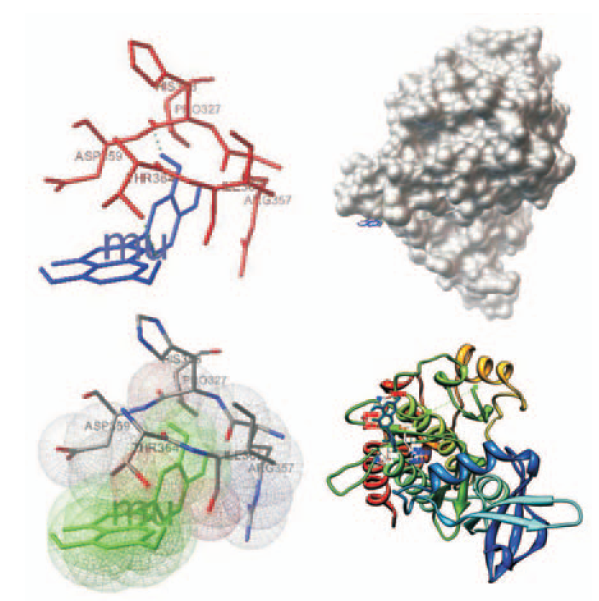
参照文献[15-16],从专业数据库获取蛋白质结构。蛋白质数据库RCSB的PDB数据库(http://www.rcsb.org/pdb/home/home.do)是一个专业收集包括蛋白质和核酸在内的生物大分子三级结构的数据库,于1971年由美国的布鲁克海文(Brookhaven)国家实验室创建。这些是构成包括细菌、酵母、植物、苍蝇、其他动物以及人类在内的生命体的基本分子。因参与肿瘤EMT过程的几大蛋白的三维晶体结构信息均可在蛋白质数据库RCSB的PDB数据库中(http://www.rcsb.org/pdb/home/home.do)上获得。从BIDD(Bioinformatics and Drug Design group)治疗性靶标数据库(Therapeutic Target Database)中收集对接受体(调控EMT的关键蛋白),包括现有常见药物靶标,结合调控EMT的几大因子,在PDB数据库搜索晶体结构,选出共20个靶标,其PDB代码见表1。对接前将配体从蛋白——配体复合物中去除后,得到各个目的蛋白的空间结构及活性口袋空间位点。通过用户图形界面ADT1.5.4版本软件中Select功能下Select from string将各个膜蛋白三维结构文件去掉水分子;同理用Select功能下Select from string窗口中Residue键入ARG8,判定对这些膜蛋白中是否含有柔性残基白;用Edit功能下Hydrogens进行加氢、合并非极性的氢,计算Gasteiger-Huekel电荷。最后以PDB格式存储,进行格式转化成PDBQT,以备Autogrid使用。
表1
Tab.1
表1
表1
蛋白PDB号、活性口袋等相关参数
Tab.1
Parameters of protein PDB number and their active pockets
目的蛋白质
PDB
X
Y
Z
长度
参考文献(第一作者、杂志名、年份)
TGF-betaR
2X7O
16.956
-4.82
2.512
342
ROTH GJ.J Med Chem,2010
EGFR
3IKA
98.27
14.08
23.68
331
ZHOU W.Nature,2009
VEGFR
2XIR
-9.236
68.598
44.359
316
IYER SJ .Biol Chem,2010
HGFR
2UZX
46.876
-34.983
130.84
289
NIEMANN HH.Cell,2007
C-Met
3R7O
90.38
100.79
-45.784
307
RICKERT KW.J Biol Chem,2011
PDGFR
1GQ4
-89.2.56
92.143
5.671
90
KARTHIKEYAN S.J Biol Chem,2002
E-cadherin
2QVF
49.341
-19.34
94.43
213
HAUSSINGER D.EMBO J,2008
N-cadherin
1NCI
24.57
56.784
100.35
110
SHAPIRO L.Nature,1995
beta-catenin
3SLA
56.498
-155.37
24.46
168
EVRARD-TODESCHI N.J Chem Inf Model,2008
IGFR
2ZM3
159.40
35.76
98.358
308
EPA VC.Protein Eng Des Sel,2006
Integrin αVβ3
1JV2
239.03
1.676
-56.140
1649
XIONG JP.Science,2001
Integrin α6β1
1AOX
98.013
-69.396
-64.13
1078
EMSLEY J.J Biol Chem,1997
Integrin αIIbβ3
3FCS
-43.654
154.93
-15.67
1649
ZHU J.Mol Cell,2008
Integrin αVβ6
3F7Q
90.256
-4.254
-67.356
896
DE PEREDA JM.EMBO J,2009
Integrin α1β1
1CK4
35.574
6.094
-57.835
198
NOLTE M.FEBS Lett,1999
Twist
1AO5
12.673
-45.670
111.56
237
TIMM DE.Protein Sci,1997
Snail
1Y62
-25.268
-178.63
9.352
160
UTSINTONG M.J Biomol Screen,2009
Slug
4MBA
79.36
-67.350
56.394
147
BOLOGNESI M.J Mol Biol,1989
Claudin-1
2QUO
100.43
-3.673
89.64
126
VAN ITALLIE CM.J Biol Chem,2008
Valentine
1PVL
67.386
-90.252
6.327
301
PEDELACQ JD.Structure,1999
Wnt
4FOA
-17.954
11.393
-13.741
316
KAKUGAWA S.Nature,2015
表1
蛋白PDB号、活性口袋等相关参数
Tab.1
Parameters of protein PDB number and their active pockets
Fig.4
Effect of luteolin on the expression of Integrin αIIbβ3 in A549 cells detected by flow cytometry A.normal control group;B.hypoxia group;C.5 μmol·L-1 luteolin group;D.10 μmol·L-1 luteolin group;E.15 μmol·L-1 luteolin group;F.25 μmol·L-1 luteolin group
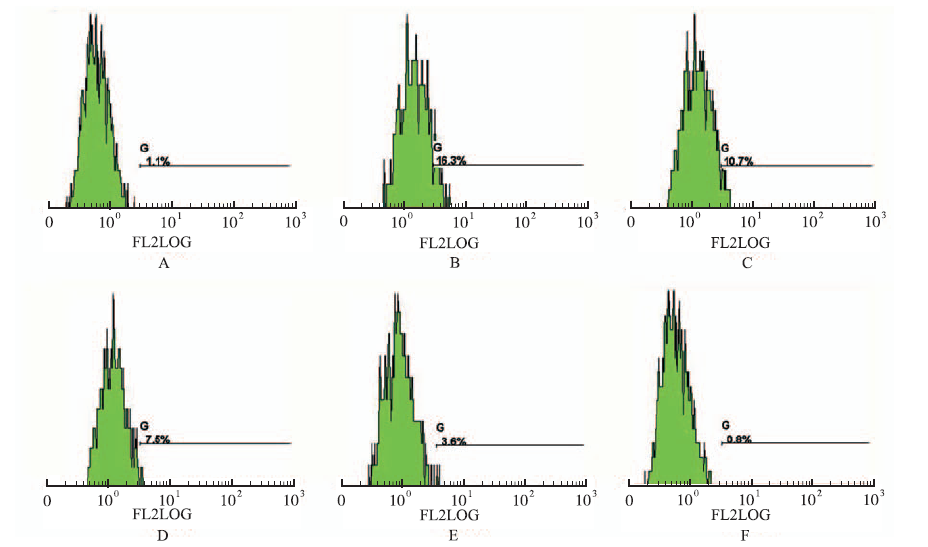
Fig.5
Effect of luteolin on the expression of Integrin αIIbβ3 in H1975 cells detected by flow cytometry A.normal control group;B.hypoxia group;C.5 μmol·L-1 luteolin group;D.10 μmol·L-1 luteolin group;E.15 μmol·L-1 luteolin group;F.25 μmol·L-1 luteolin group
TUORKEY MJ.Molecular targets of luteolin in cancer[J].Eur J Cancer Prev,2016,25(1):65-76.
Many food-derived phytochemical compounds and their derivatives represent a cornucopia of new anticancer compounds. Despite extensive study of luteolin, the literature has no information on the exact mechanisms or molecular targets through which it deters cancer progression. This review discusses existing data on luteolin anticancer activities and then offers possible explanations for and molecular targets of its cancer-preventive action. Luteolin prevents tumor development largely by inactivating several signals and transcription pathways essential for cancer cells. This review also offers insights into the molecular mechanisms and targets through which luteolin either prevents cancer or mediates cancer cell death.
SHAIKH MF,TAN KN,BORGESK.Anticonvulsant scree-ning of luteolin in four mouse seizure models[J].Neurosci Lett,2013,550(1):195-199.
Luteolin, a common plant polyphenolic flavonoid, has antioxidant, neuroprotective, anxiolytic and anti-inflammatory properties, which led us to hypothesize that luteolin is anticonvulsant. Here, we evaluated the effects of acute and chronic luteolin injection (i.p.) in four mouse seizure models, the 602Hz model, maximal electroshock test (MEST), pentylenetetrazole (PTZ) and second hit PTZ test in the chronic stage of the pilocarpine model. Using real-time PCR mRNA levels of toll like receptor 4 (Tlr4), were quantified in the pilocarpine model, because luteolin has been shown to block the downstream signaling of TLR4. Luteolin did not exhibit any consistent anti- or pro-convulsant actions after single dosing in the 602Hz (0.3–1002mg/kg), MEST (0.3–2002mg/kg) and PTZ (302mg/kg) tests, nor after repeated daily dosing (1002mg/kg) in the 602Hz model. Tlr4 mRNA levels were upregulated 3 days after pilocarpine-induced status epilepticus (SE), but unaltered at three weeks in the chronic stage of the model. At that time, there was no effect of repeated luteolin injections (1002mg/kg, i.p.) in the second hit PTZ test, indicating that TLR-4 signaling may be not one of the main players determining the seizure threshold in this seizure model. In summary, we found no indications that luteolin is pro- or anti-convulsant in one chronic and three acute mouse seizure models.
AMBASTA RK,JHA SK,KUMARD,et al.Comparative study of anti-angiogenic activities of luteolin,lectin and lupeol biomolecules[J].J Translat Med,2015,13(2):307.
Angiogenesis is a hallmark feature in the initiation, progression and growth of tumour. There are various factors for promotion of angiogenesis on one hand and on the other hand, biomolecules have been reported to inhibit cancer through anti-angiogenesis mechanism. Biomolecules, for instance, luteolin, lectin and lupeol are known to suppress cancer. This study aims to compare and evaluate the biomolecule(s) like luteolin, lupeol and lectin on CAM assay and HT-29 cell culture to understand the efficacy of these drugs. The biomolecules have been administered on CAM assay, HT-29 cell culture, cell migration assay. Furthermore, bioinformatics analysis of the identified targets of these biomolecules have been performed. Luteolin has been found to be better in inhibiting angiogenesis on CAM assay in comparison to lupeol and lectin. In line with this study when biomolecules was administered on cell migration assay via scratch assay method. We provided evidence that Luteolin was again found to be better in inhibiting HT-29 cell migration. In order to identify the target sites of luteolin for inhibition, we used software analysis for identifying the best molecular targets of luteolin. Using software analysis best target protein molecule of these biomolecules have been identified. VEGF was found to be one of the target of luteolin. Studies have found several critical point mutation in VEGF A, B and C. Hence docking analysis of all biomolecules with VEGFR have been performed. Multiple allignment result have shown that the receptors are conserved at the docking site. Therefore, it can be concluded that luteolin is not only comparatively better in inhibiting blood vessel in CAM assay, HT-29 cell proliferation and cell migration assay rather the domain of VEGFR is conserved to be targeted by luteolin, lupeol and lectin.
CHIANS,THAPAR,CHIZ,et al.Luteolin inhibits the Nrf2 signaling pathway and tumor growth in vivo[J].Biochem Biophy Res Communi,2014,447(4):602-608.
Nuclear factor erythroid 2-related factor 2 (Nrf2) is over-expressed in many types of tumor, promotes tumor growth, and confers resistance to anticancer therapy. Hence, Nrf2 is regarded as a novel therapeutic target in cancer. Previously, we reported that luteolin is a strong inhibitor of Nrf2 in vitro . Here, we showed that luteolin reduced the constitutive expression of NAD(P)H quinone oxidoreductase 1 in mouse liver in a time- and dose-dependent manner. Further, luteolin inhibited the expression of antioxidant enzymes and glutathione transferases, decreasing the reduced glutathione in the liver of wild-type mice under both constitutive and butylated hydroxyanisole-induced conditions. In contrast, such distinct responses were not detected in Nrf2 / mice. In addition, oral administration of luteolin, either alone or combined with intraperitoneal injection of the cytotoxic drug cisplatin, greatly inhibited the growth of xenograft tumors from non-small-cell lung cancer (NSCLC) cell line A549 cells grown subcutaneously in athymic nude mice. Cell proliferation, the expression of Nrf2, and antioxidant enzymes were all reduced in tumor xenograft tissues. Furthermore, luteolin enhanced the anti-cancer effect of cisplatin. Together, our findings demonstrated that luteolin inhibits the Nrf2 pathway in vivo and can serve as an adjuvant in the chemotherapy of NSCLC.
KAPOORS.Luteolin and its inhibitory effect on tumor growth in systemic malignancies[J].Experimental Cell Res,2013,319(6):777-778.
Lamy et al have provided interesting data in their recent article in your esteemed journal. Luteolin augments apoptosis in a number of systemic malignancies. Luteolin reduces tumor growth in breast carcinomas. Luteolin mediates this effect by up-regulating the expression of Bax and down-regulating the expression of Bcl-xL. EGFR-induced MAPK activation is also attenuated. As a result there is increased G2/ M phase arrest. These effects have been seen both in vivo as well as in vitro. It also reduces ERα expression and causes inhibition of IGF-1 mediated PI3K–Akt pathway. Luteolin also activates p38 resulting in nuclear translocation of the apoptosis-inducing factor. Simultaneously it also activates ERK. As a result there is increased intra-tumoral apoptosis which is caspase dependent as well as caspase independent. - Highlights: 78 Luteolin and tumor growth in breast carcinomas. 78 Luteolin and pulmonary cancer. 78 Luteolin and colon cancer.
Metastasis is the leading cause of cancer-associated death but has been difficult to study because it involves a series of rare, stochastic events. To capture these events, we developed a sensitive method to tag and track pancreatic epithelial cells in a mouse model of pancreatic cancer. Tagged cells invaded and entered the bloodstream unexpectedly early, before frank malignancy could be detected by rigorous histologic analysis; this behavior was widely associated with epithelial-to-mesenchymal transition (EMT). Circulating pancreatic cells maintaineda mesenchymal phenotype, exhibited stem cell properties, and seeded the liver. EMT and invasiveness were most abundant at inflammatory foci, and induction of pancreatitis increased the number of circulating pancreatic cells. Conversely, treatment with the immunosuppressive agent dexamethasone abolished dissemination. These results provide insight into the earliest events of cellular invasion insitu and suggest that inflammation enhances cancer progression in part by facilitating EMT and entry into the circulation.
SAKURAI MA,OZAKIY,OKUZAKID,et al.Gefitinib and luteolin cause growth arrest of human prostate cancer PC-3 cells via inhibition of cyclin G-associated kinase and induction of miR-630[J].PLoS One,2014,9(6):e100124.
Cyclin G-associated kinase (GAK), a key player in clathrin-mediated membrane trafficking, is overexpressed in various cancer cells. Here, we report that GAK expression is positively correlated with the Gleason score in surgical specimens from prostate cancer patients. Embryonic fibroblasts from knockout mice expressing a kinase-dead (KD) form of GAK showed constitutive hyper-phosphorylation of the epidermal growth factor receptor (EGFR). In addition to the well-known EGFR inhibitors gefitinib and erlotinib, the dietary flavonoid luteolin was a potent inhibitor of the Ser/Thr kinase activity of GAK in vitro. Co-administration of luteolin and gefitinib to PC-3 cells had a greater effect on cell viability than administration of either compound alone; this decrease in viability was associated with drastic down-regulation of GAK protein expression. A comprehensive microRNA array analysis revealed increased expression of miR-630 and miR-5703 following treatment of PC-3 cells with luteolin and/or gefitinib, and exogenous overexpression of miR-630 caused growth arrest of these cells. GAK appears to be essential for cell death because co-administration of gefitinib and luteolin to EGFR-deficient U2OS osteosarcoma cells also had a greater effect on cell viability than administration of either compound alone. Taken together, these findings suggest that GAK may be a new therapeutic target for prostate cancer and osteosarcoma.
TIWARIN,GHELDOFA,TATARIM,et al.EMT as the ultimate survival mechanism of cancer cells[J].Semi Can Biology,2012,22(3):194-207.
Epithelial cancers make up the vast majority of cancer types and, during the transition from benign adenoma to malignant carcinoma and metastasis, epithelial tumor cells acquire a de-differentiated, migratory and invasive behavior. This process of epithelial–mesenchymal transition (EMT) goes along with dramatic changes in cellular morphology, the loss and remodeling of cell–cell and cell–matrix adhesions, and the gain of migratory and invasive capabilities. EMT itself is a multistage process, involving a high degree of cellular plasticity and a large number of distinct genetic and epigenetic alterations, as fully differentiated epithelial cells convert into poorly differentiated, migratory and invasive mesenchymal cells. In the past years, a plethora of genes have been identified that are critical for EMT and metastasis formation. Notably, the EMT process not only induces increased cancer cell motility and invasiveness but also allows cancer cells to avoid apoptosis, anoikis, oncogene addiction, cellular, senescence and general immune defense. Notably, EMT seems to play a critical role in the generation and maintenance of cancer stem cells, highly consistent with the notion that metastatic cells carry the ability to initiate new tumors.
YEX,WEINBERG RA.Epithelial-mesenchymal plasticity:a central regulator of cancer progression[J].Trends Cell Biology,2015,25(11):675-686.
Epithelial–mesenchymal plasticity is critical for carcinoma progression and metastasis. Inhibition of EMT activation and/or epithelial–mesenchymal plasticity may serve as new ways to clinically treat advanced carcinomas.
YANGP,HUOZ,LIAOH,et al.Cancer/testis antigens trigger epithelial-mesenchymal transition and genesis of cancer stem-like cells[J].Curr Pharm Design,2015,21(10):1292-1300.
Malignant tumors aberrantly overexpress various embryonic genes and proto-oncogenes, including a variety of cancer-testis antigens (CTAs). CTAs belong to a class of testis-derived proteins which are only expressed in germ cells in the male testis, and the expression of CTA genes is entirely silenced in the adult somatic tissues. They are, however, aberrantly overexpressed in a variety of malignant tumor tissues. Emerging evidence shows that a number of CTAs promote epithelialmesenchymal transition (EMT) and genesis of cancer stem like cells, escalating tumorigenesis, invasion, and metastasis. The can cer-testis antigens, such as SSX, MAGE-D4B, CAGE, piwil2, and CT45A1, upregulate EMT and metastatic genes, promoting EMT and tumor dissemination. In addition, certain members of CTAs, including Piwil2, DNAJB8, CT45A1, MAGE-A, GAGE, and SPANX, are implicated in the initiation or maintenance, of cancer stem-like cells, promoting tumorigenesis and malignant progression. Clinically CTAs are closely associated with poor prognosis in cancer patients. Intriguely, CTAs are strongly immunogenic and normally restricted to the male testis after birth, however, these proteins are aberrantly overexpressed in cancer stem-like cells and in a variety of cancers, suggesting their target potential for cancer immunotherapy, as diagnostic biomarkers, and as targets for novel anticancer drug discovery. Thus, the targeting of tumorigenic CTAs is a promising strategy to eradicate cancer stem-like cells and inhibit tumorigenesis for effective cancer treatment.
FORLIS,HUEYR,PIQUE ME,et al.Computational prote-inligand docking and virtual drug screening with the AutoDock suite[J].Nat Protoc,2016,11(5):905-919.
Abstract Computational docking can be used to predict bound conformations and free energies of binding for small-molecule ligands to macromolecular targets. Docking is widely used for the study of biomolecular interactions and mechanisms, and it is applied to structure-based drug design. The methods are fast enough to allow virtual screening of ligand libraries containing tens of thousands of compounds. This protocol covers the docking and virtual screening methods provided by the AutoDock suite of programs, including a basic docking of a drug molecule with an anticancer target, a virtual screen of this target with a small ligand library, docking with selective receptor flexibility, active site prediction and docking with explicit hydration. The entire protocol will require 0908045 h.
UTSINTONGM,ROJSANGAP,HO KY,et al.Virtual screening against acetylcholine binding protein[J].J Biomol Screen,2012,17(2):204-215.
The nicotinic acetylcholine receptors (nAChRs) are a member of the ligand-gated ion channel family and play a key role in the transfer of information across neurological networks. The X-ray crystal structure of agonist-bound (7) acetylcholine binding protein (AChBP) has been recognized as the most appropriate template to model the ligand-binding domain of nAChR for studying the molecular mechanism of the receptor-ligand interactions. Virtual screening of the National Cancer Institute diversity set, a library of 1990 compounds with nonredundant pharmacophore profiles, using AutoDock against AChBPs revealed 51 potential candidates. In vitro radioligand competition assays using [(3)H] epibatidine against the AChBPs from the freshwater snails, Lymnaea stagnalis, and from the marine species, Aplysia californica and the mutant (AcY55W), revealed seven compounds from the list of candidates that had micromolar to nanomolar affinities for the AChBPs. Further investigation on (7)nAChR expressing in Xenopus oocytes and on the recombinant receptors with fluorescence resonance energy transfer (FRET)-based calcium sensor expressing in HEK cells showed that seven compounds were antagonists of (7)nAChR, only one compound (NSC34352) demonstrated partial agonistic effect at low dose (10 M), and two compounds (NSC36369 and NSC34352) were selective antagonists on (7)nAchR with moderate potency. These hits serve as novel templates/scaffolds for development of more potent and specific in the AChR systems.
YANJ,ZHANGG,PANJ,et al.α-Glucosidase inhibition by luteolin:kinetics,interaction and molecular docking[J].Int J Biol Macromol,2014,64(2):213-223.
α-Glucosidase is a critical associated enzyme with type 2 diabetes mellitus in humans. Inhibition of α-glucosidase is important due to the potential effect of down regulating glucose absorption in patients. In this study, the inhibitory activity of flavone luteolin on α-glucosidase and their interaction mechanism were investigated by multispectroscopic methods along with molecular docking technique. It was found that luteolin reversibly inhibited α-glucosidase in a noncompetitive manner with an IC 50 value of (1.7202±020.05)02×0210 614 02mol02L 611 , and the inhibition followed a multi-phase kinetic process with a first-order reaction. Luteolin had a strong ability to quench the intrinsic fluorescence of α-glucosidase through a static quenching procedure. The positive values of enthalpy and entropy change suggested that the binding of luteolin to α-glucosidase was driven mainly by hydrophobic interactions, and the binding distance was estimated to be 4.5602nm. Analysis of synchronous fluorescence, circular dichroism, and Fourier transform infrared spectra demonstrated that the binding of luteolin to α-glucosidase induced rearrangement and conformational changes of the enzyme. Moreover, the results obtained from molecular docking indicated that luteolin had a high affinity close to the active site pocket of α-glucosidase and indirectly inhibited the catalytic activity of the enzyme.
MARIE-EGYPTIENNE DT,LOHSEI,HILL RP.Cancer stem cells,the epithelial to mesenchymal transition (EMT) and radioresistance:potential role of hypoxia[J].Cancer Lett,2013,341(1):63-72.
Abstract Numerous studies have demonstrated the presence of cancer stem cells (CSCs) within solid tumors. Although the precursor of these cells is not clearly established, recent studies suggest that the phenotype of CSCs may be quite plastic and associated with the epithelial-to-mesenchymal transition (EMT). In patients, the presence of EMT and CSCs has been implicated in increased resistance to radiotherapy. Hypoxia, a negative prognostic factor for treatment success, is a potent driver of a multitude of molecular signalling pathways that allow cells to survive and thrive in the hostile tumor microenvironment and can induce EMT. Hypoxia also provides tumor cells with cues for maintenance of a stem-like state and may help to drive the linkage between EMT and CSCs. Understanding the biology of CSCs, the EMT phenotype and their implications in therapeutic relapse may provide crucial new approaches in the development of improved therapeutic strategies. Copyright 2012 Elsevier Ireland Ltd. All rights reserved.
Abstract Metastasis is a multi-step process which requires the conversion of polarized epithelial cells to mesenchymal cells, Epithelial-Mesenchymal Transition (EMT). EMT is essential during embryonic morphogenesis and has been implicated in the progression of primary tumors towards metastasis. Hypoxia is known to induce EMT; however the molecular mechanism is still poorly understood. Using the A431 epithelial cancer cell line, we show that cells grown under hypoxic conditions migrated faster than cells grown under normal oxygen environment. Cells grown under hypoxia showed reduced adhesion to the extracellular matrix (ECM) probably due to reduced number of Vinculin patches. Growth under hypoxic conditions also led to down regulation of E-cadherin and up regulation of vimentin expression. The increased motility of cells grown under hypoxia could be due to redistribution of Rac1 to the plasma membrane as opposed to increased expression of Rac1. EGF (Epidermal Growth Factor) is a known inducer of EMT and growth of A431 cells in the absence of oxygen led to increased expression of EGFR (EGF Receptor). Treatment of A431 cells with EGF led to reduced cell adhesion to ECM, increased cell motility and other EMT characteristics. Furthermore, this transition was blocked by the monoclonal antibody Cetuximab. Cetuximab also blocked the hypoxia-induced EMT suggesting that cell growth under hypoxic conditions led to activation of EGFR signaling and induction of EMT phenotype.
Abstract Osteosarcoma patients often exhibit pulmonary metastasis, which results in high patient mortality. Understanding the mechanisms of advanced metastasis in osteosarcoma cell is important for the targeted treatment and drug development. Our present study revealed that transforming growth factor-β (TGF-β) treatment can significantly promote the in vitro migration and invasion of human osteosarcoma MG-63 and HOS cells. The loss of epithelial characteristics E-cadherin (E-Cad) and up regulation of mesenchymal markers Vimentin (Vim) suggested TGF-β induced epithelial-mesenchymal transition (EMT) of osteosarcoma cells. TGF-β treatment obviously increased the expression of Snail, a key EMT-related transcription factor, in both MG-63 and HOS cells. Silencing of Snail markedly attenuated TGF-β induced down regulation of E-cad and up regulation of Vim. TGF-β treatment also significantly increased the expression and nuclear translocation of estrogen-related receptors alpha (ERRα), while had no obvious effect on the expression of ERα, ERβ, or ERRγ. Knock down of ERRα or its inhibitor XCT-790 significantly attenuated TFG-β induced EMT and transcription of Snail in osteosarcoma cells. Collectively, our present study revealed that TGF-β treatment can trigger the EMT of osteosarcoma cells via ERRα/Snail pathways. Our data suggested that ERRα/Snail pathways might be potential therapeutic targets of metastasis of osteosarcoma cells.
KURIMOTOR,IWASAWAS,EBATAT,et al.Drug resis-tance originating from a TGF-beta/FGF-2-driven epithelial-to-mesenchymal transition and its reversion in human lung adenocarcinoma cell lines harboring an EGFR mutation[J].Int J Oncol,2016,48(5):1825-1836.
Computational prote-inligand docking and virtual drug screening with the AutoDock suite
1
2016
... 参照文献[15-16],从专业数据库获取蛋白质结构.蛋白质数据库RCSB的PDB数据库(http://www.rcsb.org/pdb/home/home.do)是一个专业收集包括蛋白质和核酸在内的生物大分子三级结构的数据库,于1971年由美国的布鲁克海文(Brookhaven)国家实验室创建.这些是构成包括细菌、酵母、植物、苍蝇、其他动物以及人类在内的生命体的基本分子.因参与肿瘤EMT过程的几大蛋白的三维晶体结构信息均可在蛋白质数据库RCSB的PDB数据库中(http://www.rcsb.org/pdb/home/home.do)上获得.从BIDD(Bioinformatics and Drug Design group)治疗性靶标数据库(Therapeutic Target Database)中收集对接受体(调控EMT的关键蛋白),包括现有常见药物靶标,结合调控EMT的几大因子,在PDB数据库搜索晶体结构,选出共20个靶标,其PDB代码见表1.对接前将配体从蛋白——配体复合物中去除后,得到各个目的蛋白的空间结构及活性口袋空间位点.通过用户图形界面ADT1.5.4版本软件中Select功能下Select from string将各个膜蛋白三维结构文件去掉水分子;同理用Select功能下Select from string窗口中Residue键入ARG8,判定对这些膜蛋白中是否含有柔性残基白;用Edit功能下Hydrogens进行加氢、合并非极性的氢,计算Gasteiger-Huekel电荷.最后以PDB格式存储,进行格式转化成PDBQT,以备Autogrid使用. ...
Virtual screening against acetylcholine binding protein
1
2012
... 参照文献[15-16],从专业数据库获取蛋白质结构.蛋白质数据库RCSB的PDB数据库(http://www.rcsb.org/pdb/home/home.do)是一个专业收集包括蛋白质和核酸在内的生物大分子三级结构的数据库,于1971年由美国的布鲁克海文(Brookhaven)国家实验室创建.这些是构成包括细菌、酵母、植物、苍蝇、其他动物以及人类在内的生命体的基本分子.因参与肿瘤EMT过程的几大蛋白的三维晶体结构信息均可在蛋白质数据库RCSB的PDB数据库中(http://www.rcsb.org/pdb/home/home.do)上获得.从BIDD(Bioinformatics and Drug Design group)治疗性靶标数据库(Therapeutic Target Database)中收集对接受体(调控EMT的关键蛋白),包括现有常见药物靶标,结合调控EMT的几大因子,在PDB数据库搜索晶体结构,选出共20个靶标,其PDB代码见表1.对接前将配体从蛋白——配体复合物中去除后,得到各个目的蛋白的空间结构及活性口袋空间位点.通过用户图形界面ADT1.5.4版本软件中Select功能下Select from string将各个膜蛋白三维结构文件去掉水分子;同理用Select功能下Select from string窗口中Residue键入ARG8,判定对这些膜蛋白中是否含有柔性残基白;用Edit功能下Hydrogens进行加氢、合并非极性的氢,计算Gasteiger-Huekel电荷.最后以PDB格式存储,进行格式转化成PDBQT,以备Autogrid使用. ...
α-Glucosidase inhibition by luteolin:kinetics,interaction and molecular docking
Drug resis-tance originating from a TGF-beta/FGF-2-driven epithelial-to-mesenchymal transition and its reversion in human lung adenocarcinoma cell lines harboring an EGFR mutation

 , 周欢
, 周欢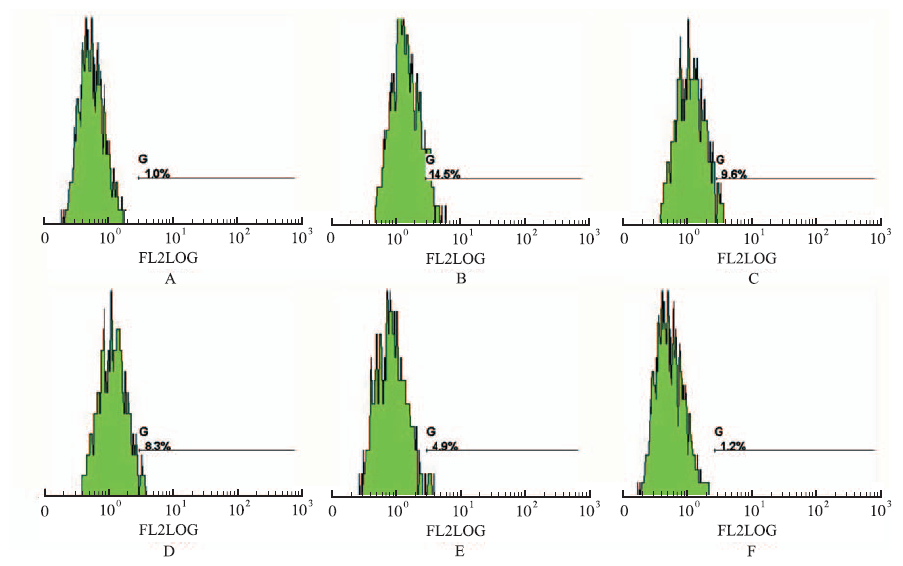


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}